As discussed in the major manuscript of this thesis, Sceloporus appear to be primitively adapted to sandy and xeric habitats [with a] nasal morphology involving an extreme lengthening of the anterior limb of the nasal passage, and the behavioral trait of "shimmy burial." The following set of Illustrations shows the range of variation in nasal morphology found within the Iguanidae.
[Note added in 2003: The relevant references are:
Stebbins, R. C. 1943. Adaptations in the nasal, passages for sand burrowing in the saurian genus Uma. Amer. Nat. 77: 38-52.
Stebbins, R. C. 1944, Some aspects of the ecology of the iguanid genus Uma. Ecol. Monogr. 14: 311-332.
Stebbins, R. C. 1948. Nasal structure in lizards with reference to olfaction and conditioning of the inspired air. Amer. J. Anat. 83: 183-222.
Stebbins performed a variety of experiments with Uma demonstrating how the anatomy of the sceloporine type nasal apparatus allowed the lizards to respire while buried in fine sand, and how the dorso-ventral flattening and musculo-sceleto system typical of most sceloporines enabled the respiration to take place by the raising and lowering of the mid thoracic body wall without loose sand filling the space required for expansion of the lungs. Stebbins also noted the sequence from a simple Anolis-like nasal apparatus, and the anatomical differences between extreme desert forms represented by Dipsosaurus (which normally respires by compressing and expanding the dorso-lateral wall of the thorax and unable to breath when buried) versus the extreme desert forms of the sceloporines that are highly adapted to respire when buried in fine sand.]
Two extremes of xeric adaptation have been found. In the Iguanidae. These are shown here by Callisaurus (Phrynosoma is a modification on the Callisaurus type probably resulting from its greatly modified skull structure) and by Dipsosaurus and Crotaphytus. Each extreme has probably evolved independently at least twice from a common mesic adapted form. Since no extreme xeric intermediate forms are known, and other intermediates clearly tend to one extreme or the other, the two extremes probably represent the developments of mutually exclusive evolutionary pathways.

Fig. 8 Crotaphytus wislisenii. Lateral and dorsal views of dissected. nasal passage, and roof of mouth (X 3.2). Transverse section at the level of the broken line, A, in the upper right hand drawing (X 5.3). [from Stebbins 1948
| a.c. | area of attachment of the concha to the lateral wall of the nasal chamber | l.w.n.c. | lateral wall of the nasal cavity |
| a.l.n.p. | anterior limb of the nasal passage | m.s.c. | median surface of the concha |
| a.s. | antiorbital space | m.w.n.c. | median wall of the nasal cavity |
| a.w.n.c. | anterior wall of the nasal cavity | n.s. | nasal septum |
| c. | concha | n.v. | nasal valve |
| c.s.m. | membrane connecting the concha with the nasal septum | np.c. | nasopharyngeal canal |
| d.l.n.p. | dorsal limb of the nasal passage | o.v.n.c. | opening of the vestibule into the primary nasal chamber |
| d.s.c. | dorsal surface of the concha | p.f. | palatine fold |
| d.w.n.c. | dorsal wall of the nasal chamber | p.l.n.p. | posterior limb of the nasal passage |
| e.n. | external naris | p.n.c. | principal nasal cavity |
| f.n.c. | floor of the nasal cavity | p.s.c. | posterior surface of the concha |
| i.n. | internal naris or choana | p.w.n.c. | posterior wall of the nasal cavity |
| l.d. | lateral diverticulum of the vestibule | s.p.n.g. | secretory pore of the lateral nasal gland |
| l.n.g. | lateral nasal gland | v. | vestibule |
| l.p. | lateral passageway between concha and lateral wall of the nasal chamber | v.l.n.p. | ventral limb of the nasal passage |
| l.p.n.s. | lateral projection from the base of the nasal septum | v.s.c. | ventral surface of the concha |
[Note added in 2003. The following illustrations are all my own, completed at the Museum of Comparative Zoology in 1972-3, based on dissections of specimens from the herpetology collection. These camera lucida drawings were done using a dissecting microscope during an exploratory survey that I intended to complete as a detailed comparative study of the evolution of the iguanid nasal aparatus. The head was skinned, the nasal, frontal, prefronal maxillary and lacrimal bones were delicately removed using microdissection tools, and the drawings were built up progressively as layers of the soft anatomy of the nasal apparatus were progressively dissected away. The Stebbins illustrations illustrate the situation of the nasal apparatus with regard to superficial anatomy. I no longer have accession numbers for the specimens illustrated.
The following illustration from this series was not bound in the thesis.
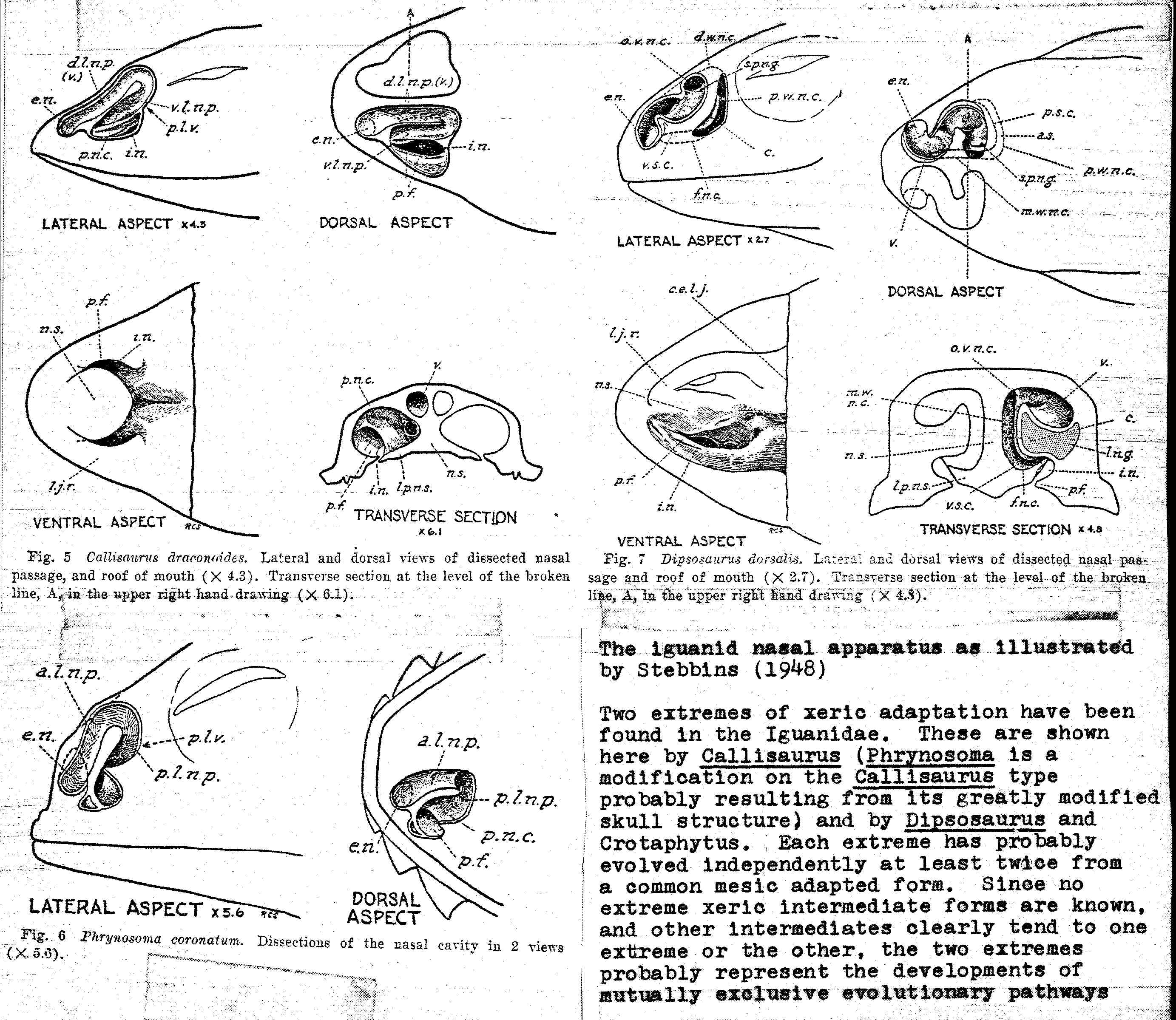
Nasal Apparatus of Anolis sagrei. The nasal morphology illustrated here is typical of all Anolis examined and has been seen in no other iguanids. Note the short, straight anterior nasal passage and relatively flat and small central chamber.]
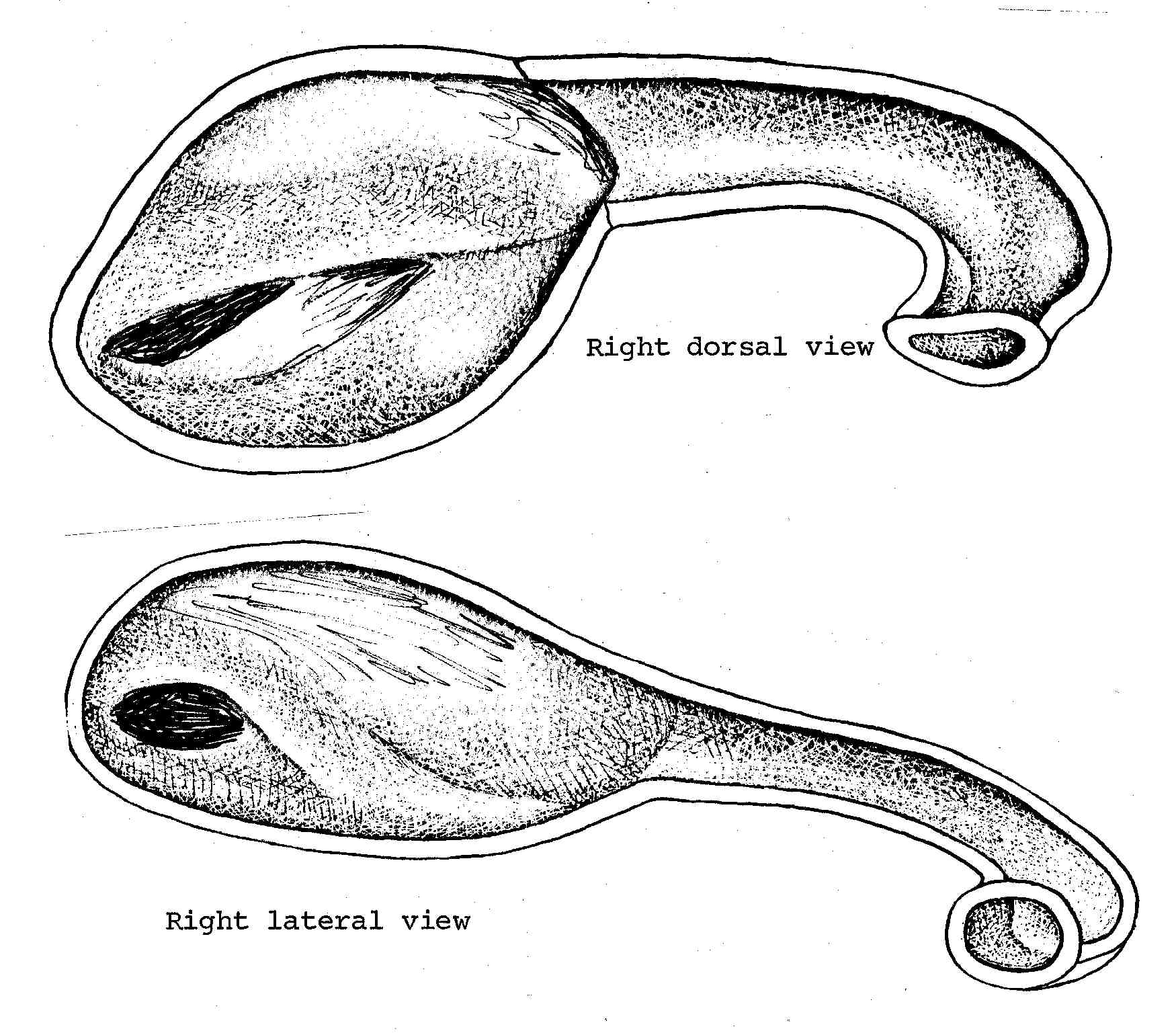
Nasal Apparatus of Leiocephalus schreibersi. The nasal morphology illustrated here is typical of Iguana and similar to that of Tropidurus torquatus, and is probably characteristic of those iguanids which have never become adapted to xeric conditions. Here the anterior nasal passage leading to the central chamber is very short and slightly curved in dorsal views. This curve may be either accentuated to increase the length of the anterior passage or it may be lengthened by growing straight back. The Anolis of the humid tropics show also an extreme reduction in the size and complexity of the nasal chamber, possibly associated with their reputed deficiency in olfaction.
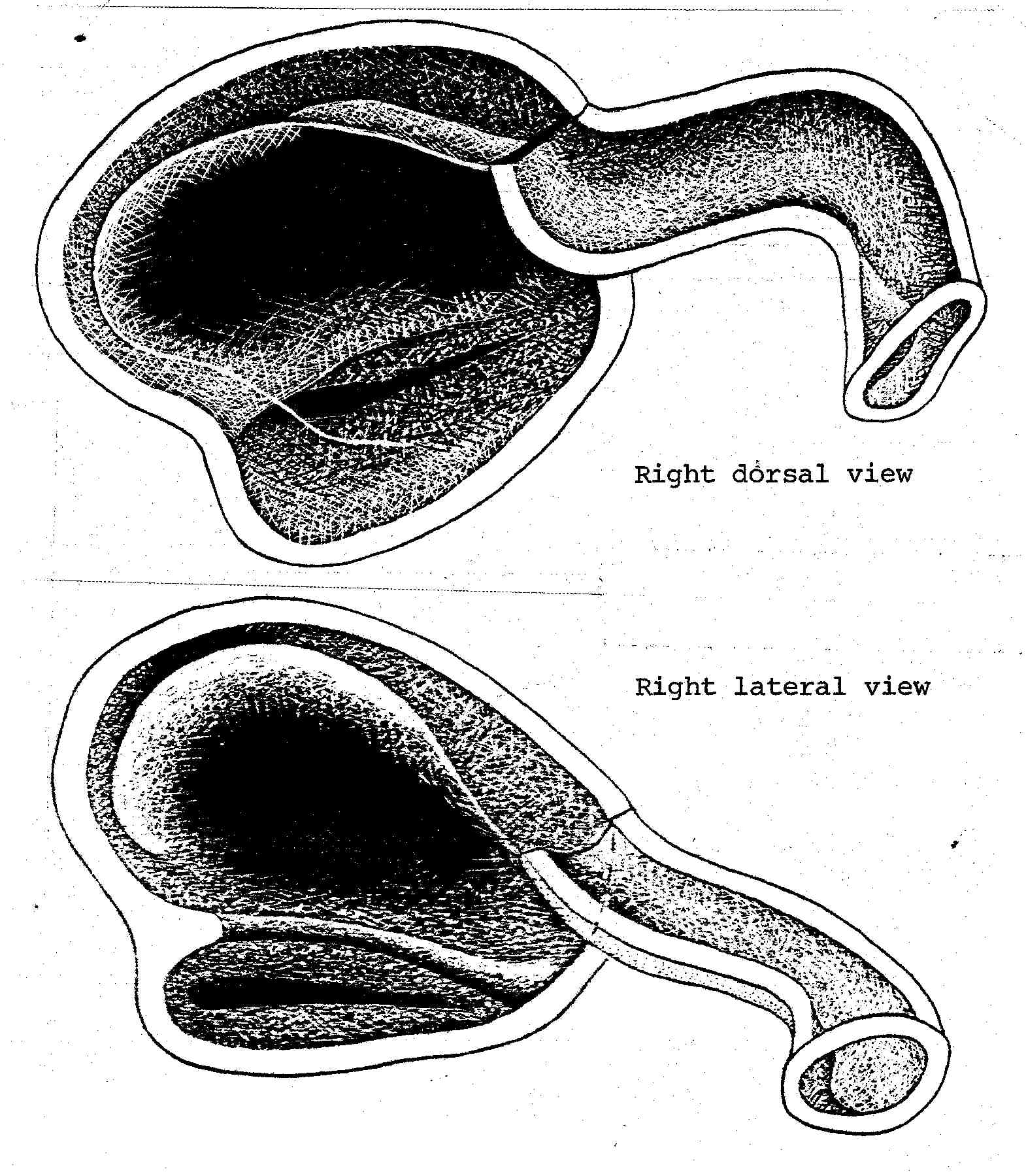
Note the extremely close parallelism with Crotaphytus and Dipsosaurus, and that Liolaemus is the major iguanid radiation in the xeric areas of southern South America. In North America the ecologically intermediate Ctenosaura show an intermediate nasal morphology, clearly on the line leading to the great sinuosity of the lengthened nasal passage seen here.
[Note from 2003: the following mss. text is associated with the original drawing above:
Nasal Apparatus of Liolaemus multiformus. The nasal morphology illustrated here is typical of Crotaphytus, Sauromalus, and Dipsosaurus. Ctenosaura is intermediate between this and the morphology illustrated for Leiocephalus. The three first mentioned genera are all adapted to the most xeric desert area. Ctenosaura extends into the margin of the desert areas. Many Liolaemus species are adapted to the deserts of Argentina and Peru.]
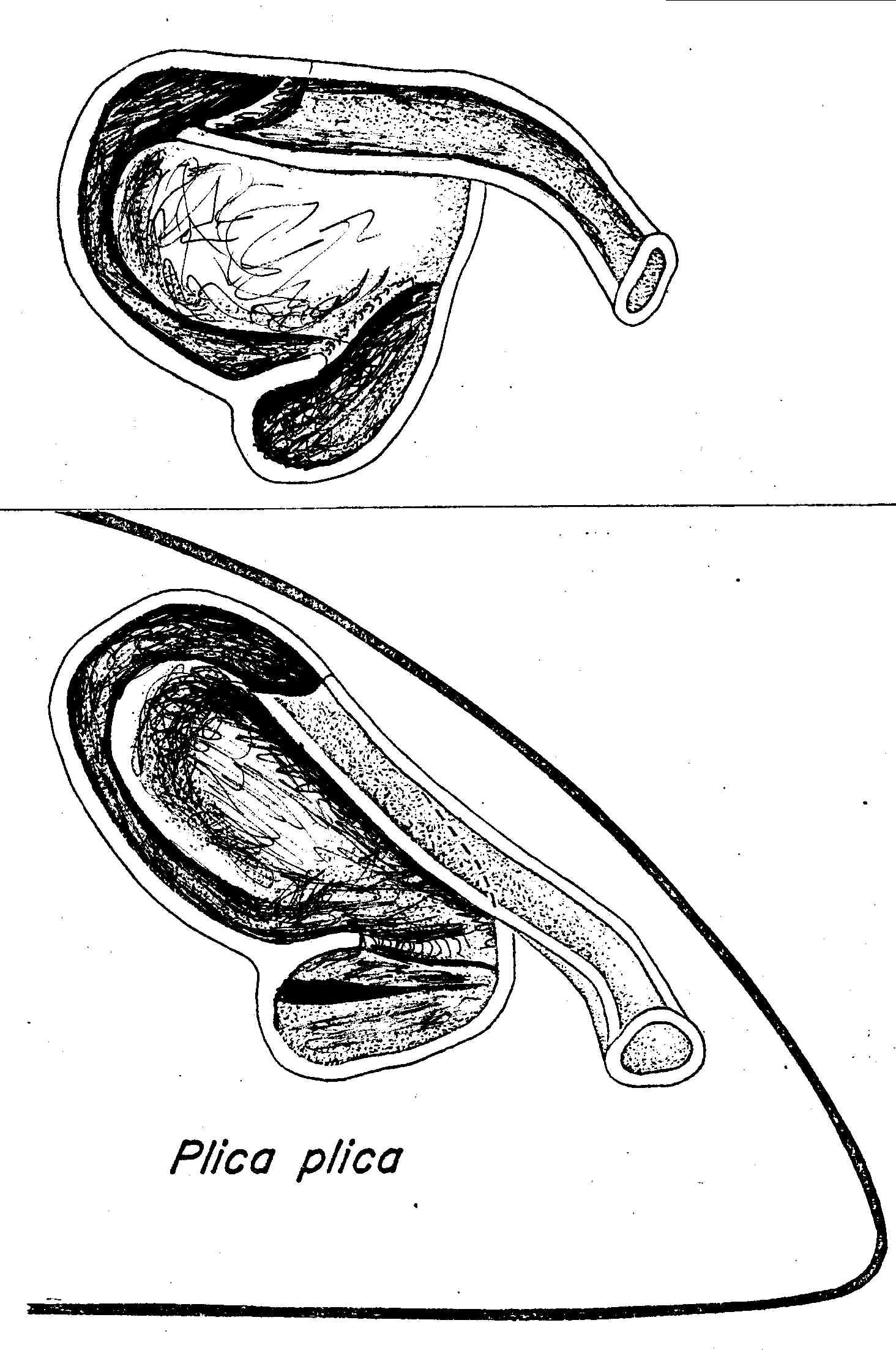
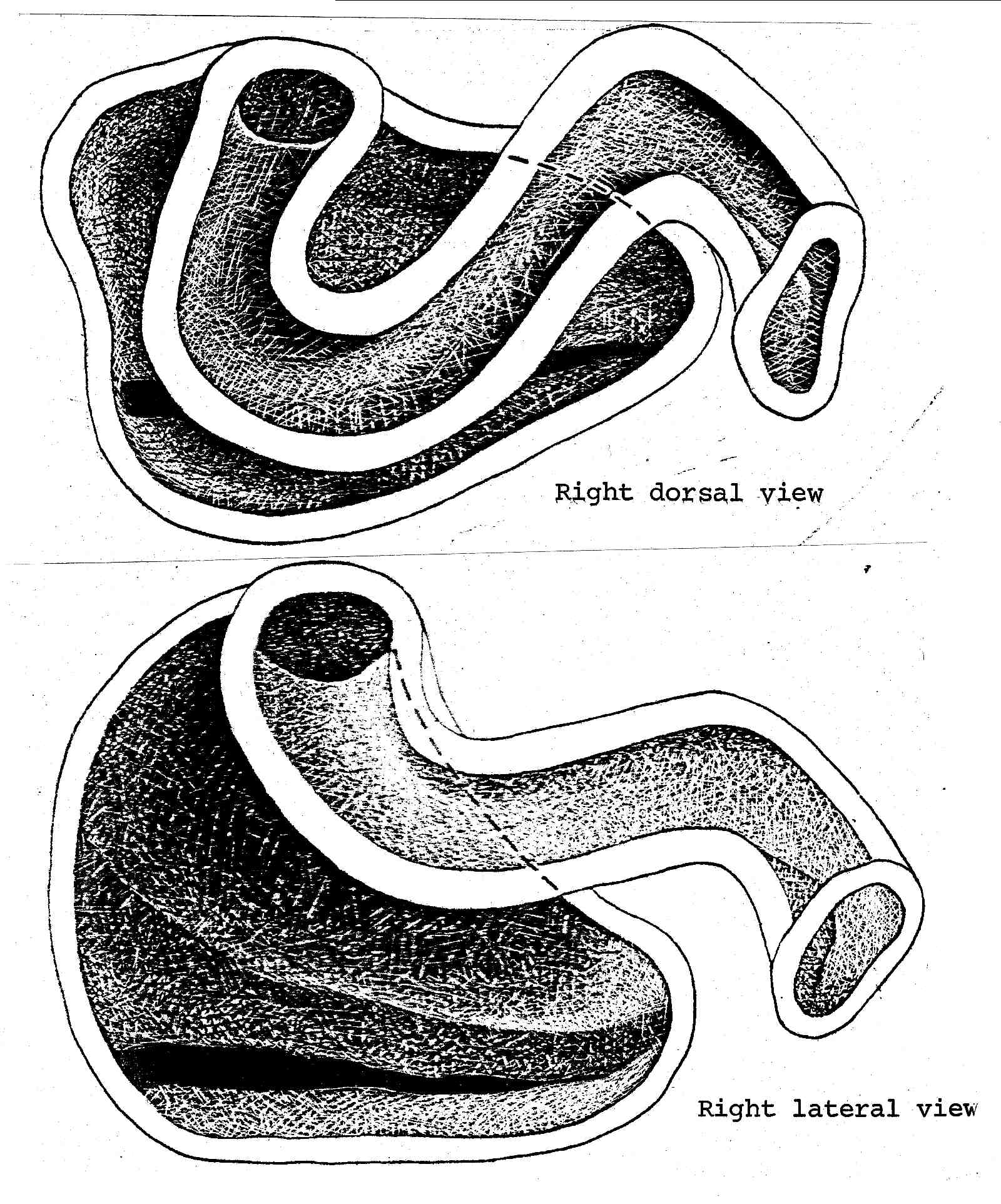
Nasal Apparatus of Plica plica. An intermediate in the evolution of the sceloporine type nasal morphology. Tropidurus torquatus shows some tendencies in this direction, and Ophryoessoides are intermediate between this and the average sceloporine condition.
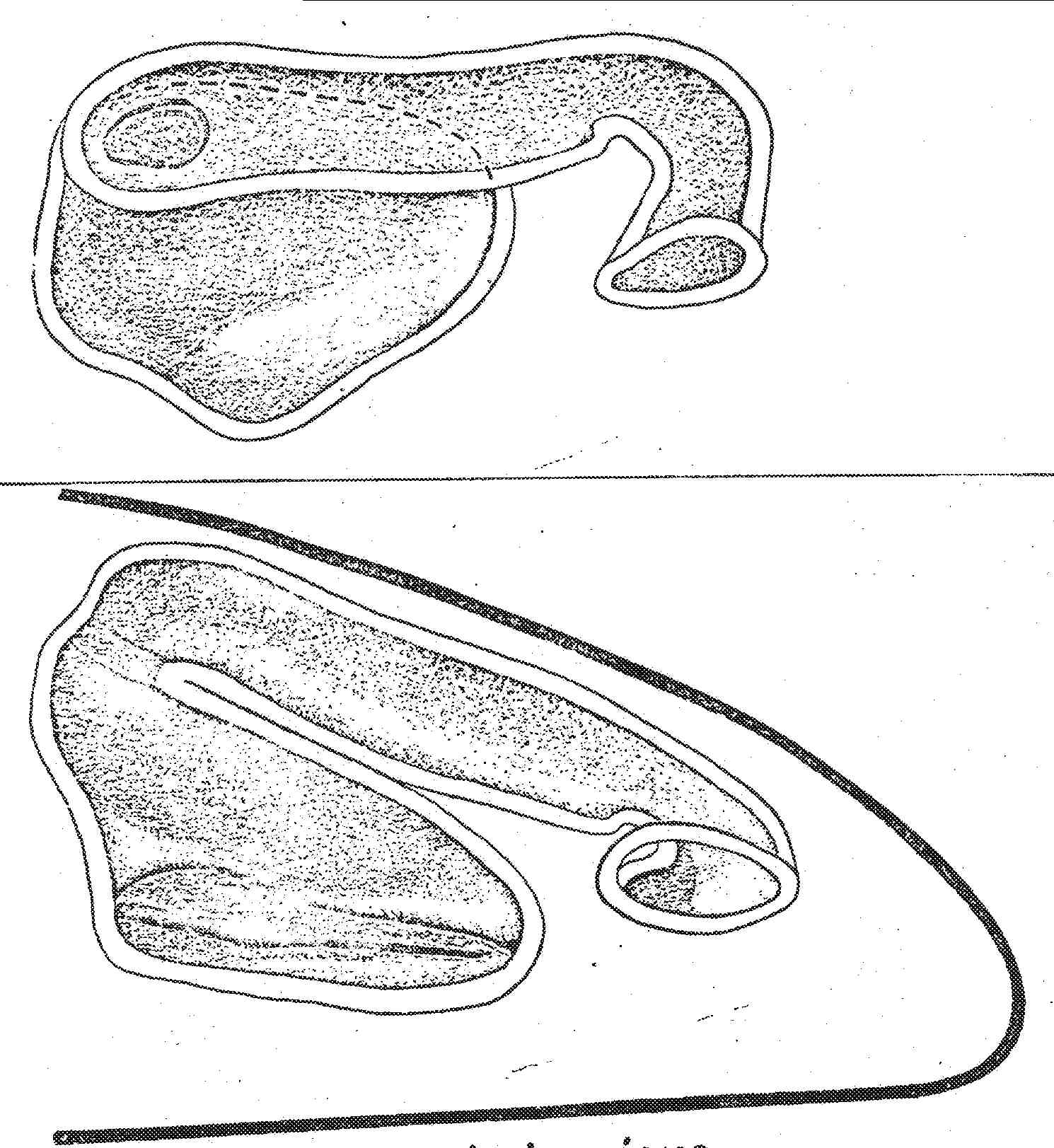
Nasal Apparatus of Petrosaurus thalassinus. Petrosaurus thalassinus has the most primitive morphology found in the sceloporines. This species is restricted to the relictual mesic areas of the sourthern end of the Baja California penninsula. Compare this with Sceloporus clarkii, illustrated on the next page, which is identical to most other sceloporines, including P. mearnsi.
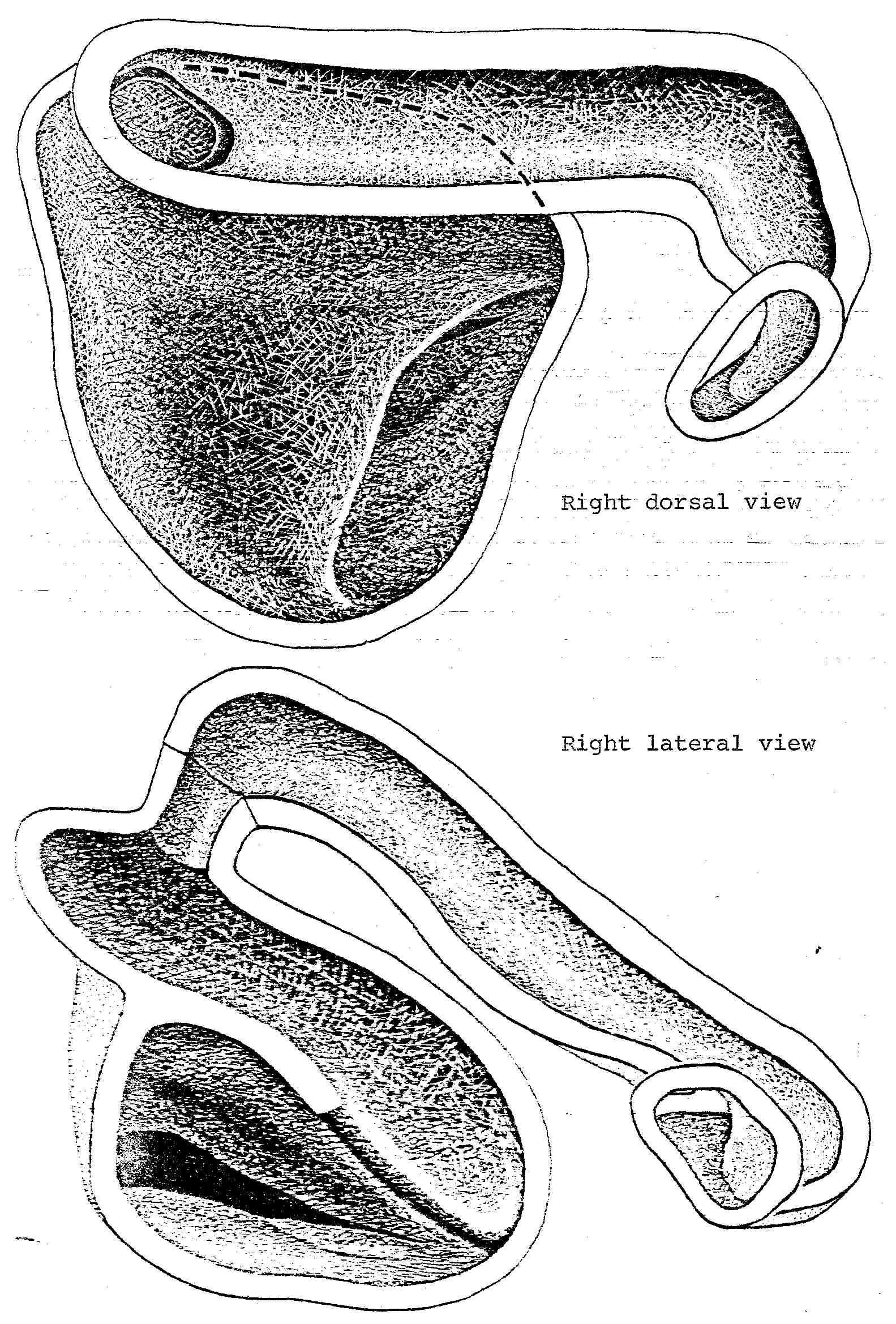
Nasal Apparatus of Sceloporus clarki. This condition is typical of P. mearnsi, Uta, Urosaurus, and all Sceloporus examined, including the tropical, arboreal, and montane formosus. A similar nasal apparatus was also found in Polychrus, at least some species of which are reported to occur in xeric areas of northern South America. Callisaurus and Uma show slightly more extreme modifications along this line, and in Phrynosoma the nasal passage is tilted so that the anterior passage rises almost vertically, but it also is cleaarly only a modification along this same line. The tropidurines Plica plica, Ophryoessoides and Petrosaurus thalassinus represent a series of intermediates between the Leiocephalus and Sceloporus condition of the nasal apparatus.